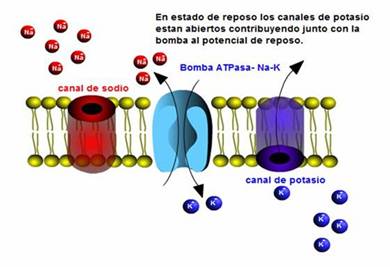
Desarrollo historico de la Biología celular.
La
teoría celular es la descripción de la constitución de los seres vivos
asentados en la estructura de las células y las aportaciones que se dieron en
la estructura de la teoría celular.
Para
esto, se encuentran diferentes estructuras respecto a la teoría celular por
medio de las aportaciones que dieron los siguientes científicos: Robert Hooke,
Anton Van Leeuwenhoek, Xavier Bichat, Theodor Schwann, Jakob Schleiden, Theodor
Boveri, Walter Sutton, Rudolf Virchow, Luis Pasteur y Santiago Ramón y Cajal.

La
aportación del científico Robert Hooke se dio utilizando su microscopio que el
mismo había fabricado. Con la
utilización de un corcho observo que se encontraba constituido por pequeñas
cavidades separadas por paredes.
Robert
Hooke nombro estas cavidades que se encontraban en el corcho con el nombre de
“CELULAS” cuyo significado es
“Habitaciones pequeñas” .
Por
lo tanto, Hooke fue el primer científico en utilizar la palabra célula para
describir los minúsculos comportamientos que constituyen un organismo.
 ¿Cómo
lo hizo? Robert Hooke dibujo 2 cortes de un trozo de corcho para observar las
paredes celulares o las células denominando que las células de estos trozos se
encuentran sin vida.
¿Cómo
lo hizo? Robert Hooke dibujo 2 cortes de un trozo de corcho para observar las
paredes celulares o las células denominando que las células de estos trozos se
encuentran sin vida.
Anton
Van Leeuwenhoek (1632-1723)
La aportación de Anton Van Leeuwenhoek se dio
utilizando el microscopio de Leeuwenhoek, se componía de una lente simple entre
dos planchas delgas de metal, en donde la muestra se colocaba en la punta de la
lanceta y esta se movia mediante un tornillo.
Fue
así que Leeuwenhoek pudo observar microorganismo vivos a través de este
microscopio. Leeuwenhoek observo en el agua estancada pequeños microorganismo
denominados “Animaculos”, que ahora denominamos Protozoarios y bacterias.
Los
protozoarios son organismos unicelulares eucariotas: heterótrofos, fagótrofos,
que viven en ambientes húmedos o directamente en medios acuáticos, ya sean
aguas saladas o aguas dulces. Leeuwenhoek también observo los glóbulos rojos de
la sangre, espermatozoides y eritrocitos humanos.


Xavier
Bichat (1771-1802)

La aportación de Xavier
Bichat, fue el que establecio que los órganos están formados por tejidos, que
forman partes diferentes de cada órgano. Los síntomas de estos órganos, derivan
la lesión de los tejidos de tal manera que estos síntomas pueden aparecer por
lesiones en órganos dieferentes.
Bitchat dio a conocer la
importancia de la patología tisular, en donde describió 21 tejidos diferentes,
que no coinciden con los actuales debido a que Bitchat no utilizo el
microscopio para describir los 21 tejidos. Por lo tanto se dice que su
aportación es extraordinaria.
En conclusión Xavier Bichat dio a conocer la
patología tisular, la cual fue su aportación básica para la teoría celular.
Theodor
Schwann (1810-1882)

La aportación de Theodor
Schwann consiste en la investigación sistemática de los tejidos animales, por
medio del origen de las células.
 Usando la microscopia
encontró el estudio de la cuerda dorsal en las larvas de rana, asi como el
cartílago. Denominando la estructura de las larva de rana, que se conforman de
células poliédricas que contiene en su interior, una formación que corresponde
como citoblasto.
Usando la microscopia
encontró el estudio de la cuerda dorsal en las larvas de rana, asi como el
cartílago. Denominando la estructura de las larva de rana, que se conforman de
células poliédricas que contiene en su interior, una formación que corresponde
como citoblasto.
La
estructura de los tejidos vegetales se debe también al cartílago, por tanto
descubrió que en una y otro la formación de nuevas células a partir de núcleos.
Por tanto, distinguió las
células de la cuerda dorsal y del cartílago se derivan de estructuras idénticas
a las células vegetales con nucleo, membrana y vacuola. Como también afirma y
confirma la primera teoría celular, a través de la estructura unitaria y común
de los seres vivos, sean animales o vegetales por medio de las células.
En conclusión, Theodor
Schwan denomino el Reino Animal y Vegetal a través de la estructura de la
celula.
Matthias
Jakob Schleiden (1804-1881)

La aportación de Jakob
Schleiden consiste en todos los organismo están constituidos de acuerdo con un
mismo principio general, y este principio es la formación de células, por tanto
toda planta está hecha de células al igual que todo animal esta hecho de
células.
Estudio el crecimiento de
las plantas por medio de las células en donde se limitaba a nombrar y describir
las plantas.
Con la utilización del
microscopio propuso que las plantas estaban compuestas por células, por lo
tanto obtuvo que el crecimiento de las plantas se producia por medio de células
nuevas que se propagan a partir de los nucleos en las células viejas.
Theodor
Boveri (1862–1915)

La
aportación de Theodor Boveri fue la propuesta de la Teoría Cromosómica de la
herencia.
Boveri clarifico que todas
las células provenientes por división de un huevo fecundado, la mitad de los
cromosomas son de origen paterno y la otra mitad es de origen materno.
Por medio de la división,
las células hijas tienen de nuevo el mismo orden cromosomatico y de esta manera
Theodor Boveri estableció la teoría de la individualidad de los cromosomas.
Boveri había encontrando la
meiosis, es
la división celular que permite la reproducción sexual. Comprende dos
divisiones sucesivas: una primera división meiótica, de una célula madre
diploide (2n) se obtienen dos células hijas haploides (n). Segunda división
meiótica, las células hijas tienen el mismo número de cromosomas que la célula
madre (como la división mitótica).
Walter
Sutton (1877-1916)

La
aportación de Walter Sutton fue la
teoría de que las leyes mendelianas de la herencia podían ser aplicadas a los
cromosomas a nivel celular. Por lo tanto propuso que los genes se localizaban
en los cromosomas. Entre Theodor Boveri y Walter Sutton propusieron la Teoría
Cromosómica de la herencia.
Se
baso es los factores hereditarios de Mendel porque decía que ahí se encontraban
los cromosomas porque creían que la separación de los cromosomas durante la
meiosis es la basa para explicar las
leyes de Mendel.
En conclusión Sutton,
relaciono las leyes de Mendel con el comportamiento de los cromosomas durante la meiosis y la fecundación

Rudolf Virchow
(1821-192)
La
aportación de Rudolf Virchow fue la patología celular donde explicó que toda
célula se ha originado a partir de otra célula, por medio de la división.
Virchow
dio la tercera preposición de la célula, con base de las investigaciones en el
campo de la citología dijo que:
Toda
célula proviene de otra célula viva semejante preexistente. Por lo cual integro
finalmente el concepto universal de célula, por lo cual se afirma que la célula
es la unidad anatómica y fisiológica que da origen a los seres vivos.
 Louis Pasteur (1822-1895)
Louis Pasteur (1822-1895)
La
aportación de Louis Pasteur fue la idea de que un organismo vivo no surgen en
forman espontanea sino que pueden ser generados solo por organismo
preexistentes fue activamente defendida y confirmada por Pasteur.
Por
lo tanto Pasteur postulo la teoría microbiana donde encontramos los
antibióticos, la anestesia, la inmunología y enfermedades.
Por
medio de la teoría microbiana, Pasteur tomo la idea de acción nociva de agentes
biológicos para la fabricación de vacunas.
El
origen de la inmunología se identifica con el de vacunas y el primer método
general para reproducirlas.

Santiago Ramón y Cajal
La aportación
de Santiago Ramón y Cajal fue la
extensiva al sistema nervioso de la teoría celular, por lo tanto denomino la
“Teoría de la neurona”.
Como
se dio esta teoría, el sistema nervioso se encuentra formado por células
conectadas entre si por sus propias prolongaciones.



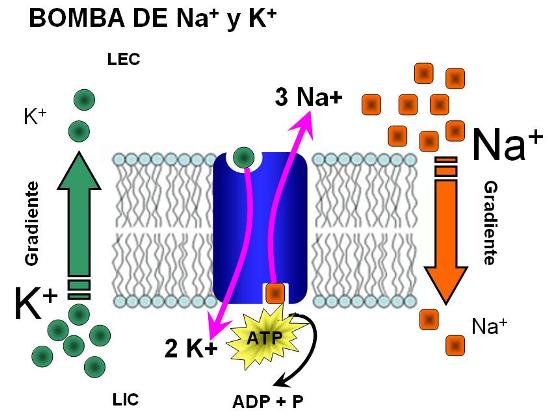
![[difusion.gif]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEhQsxRvayH9FyCm1inLK1PfmZPMKyhYubKYwFfU83Ac6OU_7TEn79GP5fLkzcX75k_s8_1fm8J_Ypq4uCh3dqFTnHCMaSCOO8X-ej8i3Mk0ZJwV-t4IXVi_Fk-jZQ5ZH9IEtlVLKQPa7ZU/s200/difusion.gif)






![[Exocitosis.jpg]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEjsTTVBztTj-gUywYtebbq4PZf7y8LC4wdOV6l_Id5lmYCMfrWH2LAB088do0ty-BhxUSqVQzahZZUo5Nmie9ESlI5Tz3kyxEpvqsJkZhKsaPOcxNzX-mcJfGyeEFwcDPs9Sh0jZVljNgA/s400/Exocitosis.jpg)