II. ESTRUCTURA DE LA MEMBRANA NUCLEAR
Cada célula por una membrana celular. La membrana celular forma una barrera permeable selectiva entre el citoplasma y el medio externo.
Composición molecular:
la membrana plasmática se compone de una bicapa fosfolipidica y proteínas integrales y periféricas relacionadas.
Está compuesta por una serie de
sustancias entre las que se destacan los lípidos, las proteínas, los glúcidos y
el colesterol (un tipo de lípido).Se discutió y estudió por mucho tiempo para
poder comprender como, los compuestos mencionados se organizan para formar la
membrana. Se postularon diversas teorías, una de ellas, la más aceptada por los
científicos es el modelo de mosaico fluido
• Modelo: porque no está
demostrada en un cien por cien, aún quedan ciertos puntos oscuros en su
organización, pero sirve para explicar varias características de la membrana.
• Mosaico: porque tiene muchos
componentes (lípidos, proteínas, glúcidos, colesterol)
• Fluido: porque los componentes
no están fijos, se mueven. Presenta características propias de los líquidos.
actúa para:
- conservar la integridad estructural de la célula
- controlar movimientos de sustancias hacia el interior y exterior de la célula (permeabilidad selectiva)
- regular interacciones entre las células
- reconocer, mediante receptores, antígenos y células extrañas así como células alteradas
- actuar como una interfaz entre el citoplasma y el medio externo
- establecer sistemas de transporte para moléculas especificas
- transferir señales físicas o químicas extracelulares a fenómenos intracelulares

TRANSPORTE DE MOLECULAS A TRAVES DE LA MEMBRANA
De todas las propiedades
descritas en el modelo que tienen las membranas, se desprende una que es la más
relevante desde el punto de vista funcional: La permeabilidad selectiva, es
decir, la posibilidad de que la membrana restrinja los solutos que han de pasar
a su través, pudiendo variar dicha permeabilidad en función de las necesidades
celulares en cada momento.
- Difusión simple
 Es el movimiento de las
partículas desde un área donde la concentración que presenta es alta, a un área
con baja concentración
Es el movimiento de las
partículas desde un área donde la concentración que presenta es alta, a un área
con baja concentración
Las moléculas pequeñas y no
polares, atraviesan la bicapa lipídica por difusión simple. Durante la
difusión, las moléculas tienden a atravesar la membrana desde regiones de alta
concentración hacia regiones de baja concentración, hasta alcanzar el
equilibrio: igual proporción de moléculas atravesarán la membrana en ambas
direcciones.
![[difusion.gif]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEhQsxRvayH9FyCm1inLK1PfmZPMKyhYubKYwFfU83Ac6OU_7TEn79GP5fLkzcX75k_s8_1fm8J_Ypq4uCh3dqFTnHCMaSCOO8X-ej8i3Mk0ZJwV-t4IXVi_Fk-jZQ5ZH9IEtlVLKQPa7ZU/s200/difusion.gif)
- Difusión facilitada
Cuando las moléculas son
impulsadas por un gradiente de concentración, el paso de la mayor parte de los
iones y moléculas a través de una pequeña membrana requiere la ayuda de
proteínas de transporte de membrana, sea proteínas del canal o proteínas
transportadoras

La difusión facilitada utiliza
canales (formados por proteínas de membrana) para permitir que moléculas
cargadas (que de otra manera no podrían atravesar la membrana) difundan
libremente hacia afuera y adentro de la célula. Estos canales son usados sobre
todo por iones pequeños tales como K+, Na+, Cl-.
La velocidad del transporte
facilitado está limitado por el número de canales disponibles mientras que la velocidad
de difusión depende solo del gradiente de concentración.
- Osmosis
La ósmosis es una clase especial
de difusión que se define como "el flujo neto de agua que atraviesa una
membrana semipermeable que separa dos compartimentos acuosos".
 La membrana celular se comporta
de forma aproximada como una membrana semipermeable, es decir dejando pasar el
agua pero no los solutos. El agua se mueve desde una zona donde su
concentración es mayor, a otra donde es menor. En el caso de las soluciones
intra y extracelular el agua se moverá desde la solución que presente una menor
concentración (solución hiposmótica) a la que tenga la mayor concentración
(solución hiperosmótica). La presión hidrostática necesaria para impedir la
ósmosis se define como "presión osmótica".
La membrana celular se comporta
de forma aproximada como una membrana semipermeable, es decir dejando pasar el
agua pero no los solutos. El agua se mueve desde una zona donde su
concentración es mayor, a otra donde es menor. En el caso de las soluciones
intra y extracelular el agua se moverá desde la solución que presente una menor
concentración (solución hiposmótica) a la que tenga la mayor concentración
(solución hiperosmótica). La presión hidrostática necesaria para impedir la
ósmosis se define como "presión osmótica".
La osmolaridad total de una
solución es la suma de la osmolaridad de cada uno de sus solutos
constituyentes. Los líquidos corporales tienen una osmolaridad de 0,29
osmoles/litro o 290 mO/l. Los iones principales (Na, K, Cl, etc.) son
responsables del 96%, mientras que la glucosa, aminoácidos y otros solutos no
disociables aportan tan sólo un 3%; de estos solutos, las proteínas tan sólo
constituyen el 0,5% de la osmolaridad total del plasma y valores aún meno- res
en los líquidos extracelulares, prácticamente carentes de proteínas. A pesar de
su pequeña proporción, la presión osmótica de las proteínas plasmáticas,
denominada presión coloidosmótica o presión oncótica es muy importante en el
intercambio de líquidos en el capilar
Son proteínas integrales
que usan gradientes electroquímicos para mover
moléculas entre ambos lados de la membrana. Este tipo de movimiento se
denomina difusión facilitada: difusión porque es un movimiento pasivo generado
por el gradiente electroquímico existente y facilitada puesto que es el
transportador el que la permite. Los transportadores son muy numerosos, más de
100 familias, y aparecen en todas las membranas de la célula. El mecanismo de
transporte supone un reconocimiento de la molécula o moléculas a las que van a
transportar y un cambio conformacional del transportador que posibilita el
trasiego de las moléculas entre ambos lados de la membrana.
Puede ser de distintos
tipos:
· Uniporte
El transporte uniporte
supone mover una molécula a favor de su gradiente de concentración
·
Contraporte
El cotransporte
permite la translación simultánea de dos moléculas entre ambos lados de la
membrana.
·
simporte
Si las dos moléculas viajan en el mismo sentido se denomina simporte.
·
Antiporte
Si el sentido en el que viaja una molécula es
contrario al de la otra se denomina antiporte

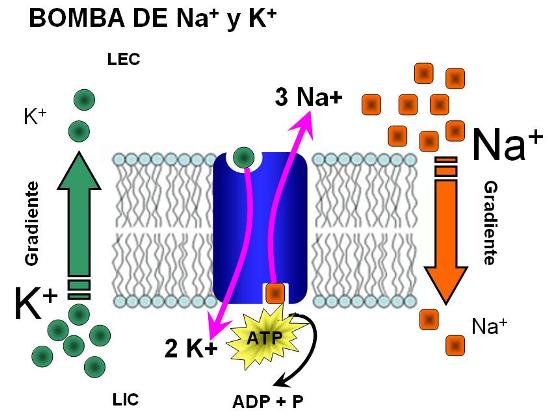
La bomba Na+K+-ATPasa
La bomba sodio-potasio consta de
dos proteínas globulares distintas: una de mayor tamaño denominada subunidad
alfa, que tiene un peso molecular de aproximadamente 100.000, y una más pequeña
denominada subunidad beta, que tiene un peso molecular de aproximadamente
55.000.
El funcionamiento de la bomba es
el siguiente: cuando dos iones potasio se unen al exterior de la proteína
transportadora y tres iones sodio se unen al interior se activa la función
ATPasa de la proteína la cual escinde una molécula de ATP, dividiéndola en ADP
y liberando un enlace de energía de fosfato de alta energía. Se cree que esta
energía liberada produce un cambio químico y conformacional en la molécula
transportadora proteica, transportando los tres iones sodio hacia el exterior y
los dos iones potasio hacia el interior
Una de las funciones más
importantes de la bomba sodio-potasio es controlar el volumen de todas las
células. Sin la función de esta bomba la mayor parte de las células del cuerpo
se hincharía hasta explotar
La bomba Ca++-ATPasa
Otra bomba importante en la
función celular es la bomba Ca++-ATPasa que se encarga de mantener los niveles
intracelulares de calcio a un nivel bajo. En algunas células, como los
eritrocitos, está localizada en la membrana celular y su función es transportar
Ca++ fuera de la célula. Sin embargo, en las células musculares, la bomba
Ca++-ATPasa se encuentra en la membrana del retículo sarcoplásmico. La bomba
transporta el Ca++ desde el citosol hacia el interior del orgánulo, que
concentra y almacena el calcio. La salida del Ca++ del retículo sarcoplásmico
al citosol muscular origina la contracción de la célula y se requiere una
rápida eliminación de este calcio para que la célula se relaje.
La bomba Ca++-ATPasa funciona de una forma muy
parecida a la bomba Na+K+-ATPasa: dos iones de Ca++ son transportados fuera del
citosol por cada molécula de ATP hidrolizado. La actividad de esta bomba está
regulada de tal forma que si la concentración de Ca++ aumenta, la velocidad de
bombeo aumenta hasta que la concentración citosólica se reduce a 0.1
mmolar.
TANSPORTE DE MACROMOLECULAS
Las macromoléculas y las grandes partículas se incorporan y se eliminan de la célula mediante procesos en los que se produce una deformación de la membrana plasmática.
El proceso de incorporación recibe el nombre de endocitosis, mientras que el de eliminación se llama exocitosis
El proceso de incorporación recibe el nombre de endocitosis, mientras que el de eliminación se llama exocitosis
LA ENDOCITOSIS. Es el proceso mediante el cual se incorporan sustancias a la célula. De esta manera, las sustancias son transportadas al interior celular a través de pequeñas depresiones de la membrana plasmática, que llevan a la formación de una vesícula endocítica. Se distinguen tres tipos de endocitosis:
- Fagocitosis
Durante este proceso, la célula genera proyecciones de la membrana y del citoplasma llamadas pseudópodos, que rodean a una partícula sólida. Una vez rodeada, los pseudópodos se fusionan formando una vesícula alrededor de la partícula llamada vesícula endosómica, la que al fusionarse con lisosomas forma la vesícula fagosómica. Estas vesículas alcanzan un tamaño mayor de 150 nm de diámetro y en su interior contienen restos celulares, microorganismos o partículas de gran tamaño. El material sólido al interior de la vesícula es digerido por las enzimas lisosomales. Los glóbulos blancos son el principal ejemplo de células que fagocitan bacterias y otras sustancias extrañas, proceso que forma parte del mecanismo de defensa del organismo.

- Pinocitosis
En este proceso, la sustancia transportada es un fluido de líquido extracelular. En este caso, no se forman pseudopodos, sino que la membrana se repliega o se invagina creando una vesícula pinocítica, con un diámetro menor a 150 nm. Una vez que el contenido de la vesícula ha sido procesado por parte de la célula, la membrana que compone la vesícula retoma a la superficie de la célula. De este modo, hay un tránsito constante de membranas entre la superficie de la célula y su interior.

- Endocitosis mediada por receptor
Este proceso es similar a la pinocitosis; sin embargo, presenta algunas diferencias. En este proceso, la invaginación de la membrana tiene lugar cuando una molécula determinada, llamada ligando, se une a un receptor ubicado en la membrana, vale decir, a una proteína específica que reconoce a un ligando específico, existente en la membrana.
Una vez formada la vesícula endocítica, esta se une a otras vesículas para formar una estructura mayor llamada endosoma. Dentro del endosoma se produce la separación del ligando y del receptor. De esta forma, los receptores son liberados y devueltos a la membrana, mientras que el ligando se fusiona con un lisosoma, siendo digerido por las enzimas de este último.

![[Exocitosis.jpg]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEjsTTVBztTj-gUywYtebbq4PZf7y8LC4wdOV6l_Id5lmYCMfrWH2LAB088do0ty-BhxUSqVQzahZZUo5Nmie9ESlI5Tz3kyxEpvqsJkZhKsaPOcxNzX-mcJfGyeEFwcDPs9Sh0jZVljNgA/s400/Exocitosis.jpg) LA EXOCITOSIS. Es el proceso contrario a la endocitosis. Mediante este proceso vesículas intracelulares se fusionan con la membrana plasmática y liberan al exterior su contenido. De esta forma las células liberan macromoléculas sintetizadas por ellas y productos de desecho. Mediante la exocitosis la membrana de la vesícula intracelular se incorpora a la membrana plasmática aumentando la superficie celular, mientras que con la endocitosis ocurre lo contrario. Por lo que es necesario que haya equilibrio entre ambos procesos para mantener invariable el volumen celular.
LA EXOCITOSIS. Es el proceso contrario a la endocitosis. Mediante este proceso vesículas intracelulares se fusionan con la membrana plasmática y liberan al exterior su contenido. De esta forma las células liberan macromoléculas sintetizadas por ellas y productos de desecho. Mediante la exocitosis la membrana de la vesícula intracelular se incorpora a la membrana plasmática aumentando la superficie celular, mientras que con la endocitosis ocurre lo contrario. Por lo que es necesario que haya equilibrio entre ambos procesos para mantener invariable el volumen celular.
POTENCIAL DE MEMBRANA
El potencial de membrana es el resultado de la separación de cargas positivas y negativas a través de una membrana celular. Esta separación, cargas positivas en el exterior de la membrana de una célula del sistema nervioso en reposo, es posible debido a que la bi-capa lipídica actúa como una barrera para la difusión de los iones y da lugar a la generación de una diferencia de potencial.
El potencial
de membrana es el voltaje que le dan a la membrana las concentraciones de los
iones en ambos lados de ella

EQUILIBRIO IÓNICO
Dado que los canales iónicos reconocen y seleccionan iones específicos, la distribución de especies iónicas a través de la membrana dependerá de la distribución particular de canales iónicos que tenga la membrana celular.
Las especies iónicas no se distribuyen de igual manera en ambos lados de la membrana:
- El sodio y el cloruro están más concentrados en el exterior de la célula
- El potasio y los aniones orgánicos (ácidos orgánicos y proteínas) están más concentrados en el interior.
Analizando por ejemplo al ión cloruro, ya que se encuentra más concentrado en el exterior de la célula tendrá la tendencia a moverse hacia el interior a través de los canales “nongated”. Sin embargo si se tiene en cuenta que en el interior de la célula existe un exceso de carga negativa esto provocará la salida de los iones cloruros.
De lo anterior se espera se alcance un estado de equilibrio, directamente relacionado con el potencial de membrana.
Para el caso de los cationes, se debe hacer el análisis conjunto del movimiento de los iones sodio y potasio, ya que el movimiento de los iones sodio hacia el interior de la célula tiene que ser compensado por la salida de iones potasio.
Este intercambio estacionario de iones previene una despolarización reversible, sin embargo no puede sostenerse indefinidamente a menos que se revierta el proceso. Esto se consigue mediante la acción de las “bombas de sodio y potasio”. Estos sistemas permiten la salida del sodio del interior de la célula e incorporan el potasio.
Este proceso implica un consumo de energía puesto que los movimientos se realizan en contra de los gradientes electroquímicos. Esta energía se obtiene de la hidrólisis de ATP, que es la molécula “portadora de energía” por excelencia. Algunas células poseen también “bombas de cloro” que funcionan de la misma manera que la mencionada arriba.
POTENCIAL DE MEMBRANA EN REPOSO
Cuando una célula está en reposo, el potencial de membrana se conoce como potencial de reposo. Por convención se toma el potencial externo como cero, por lo que teniendo en cuenta que el interior tiene un exceso de carga negativa, el potencial de membrana en este caso toma valor negativo de -60 a -70 mV.
La separación de carga a través de la membrana, y por lo tanto el potencial de reposo se modifica cada vez que ocurre un flujo neto de iones hacia o desde el interior de la célula. Cuando se produce una disminución de la separación de carga el proceso se denomina depolarización. El caso contrario se conoce como hiperpolarización.
depende de:a) La redistribución de los iones a través de la membrana. El Na+ y Ca+2 más abundantes afuera. El K+, PO4-, SO4- y Cl- más abundante adentro.
b) La acción de la bomba Na-K ATPasa. La bomba saca 3 sodios e introduce 2 potasios simultáneamente, manteniendo una carga positiva afuera.
c) Los canales de K de fuga siempre abiertos. El potasio sale por los canales abiertos atraído por el gradiente químico y luego incrementa la repulsión eléctrica con los iones de sodio, incrementándose el gradiente eléctrico que repele al potasio. Se crea un equilibrio entre ambos gradientes y el potasio deja de salir quedándose más concentrado en el interior de la célula.
b) La acción de la bomba Na-K ATPasa. La bomba saca 3 sodios e introduce 2 potasios simultáneamente, manteniendo una carga positiva afuera.
c) Los canales de K de fuga siempre abiertos. El potasio sale por los canales abiertos atraído por el gradiente químico y luego incrementa la repulsión eléctrica con los iones de sodio, incrementándose el gradiente eléctrico que repele al potasio. Se crea un equilibrio entre ambos gradientes y el potasio deja de salir quedándose más concentrado en el interior de la célula.
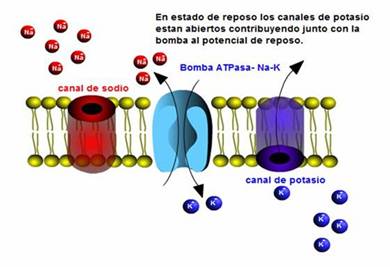
POTENCIAL DE ACCIÓN
Así como el potencial de reposo se corresponde con un estado donde la neurona está sin estimular, cuando es estimulada se produce una “explosión de actividad eléctrica” conocido como potencial de acción.
Ante el estímulo, el potencial de reposo toma valores más positivos, pero solo cuando alcanza un valor umbral, de unos – 55 mV, se produce el disparo de un potencial de acción
En términos generales el proceso puede describirse teniendo en cuenta el funcionamiento de los canales iónicos. Ante el estímulo se abren primero los canales de sodio permitiendo el ingreso al interior de la célula lo que conduce a la depolarización. Cuando los canales de sodio comienzan a cerrarse se abren los de potasio, permitiendo la salida del mismo con lo que se revierte la depolarización anterior. Con esto se regresa al valor de potencial inicial (-70mV) es decir ocurre la repolarización.

REFERENCIAS
·
Ángel Luis García Villalón, transporte de membrana [en
línea] Departamento De Fisiología Facultad De Medicina Universidad autónoma de
Madrid [fecha de acceso: 14 de septiembre de 2014]. URL disponible en:
http://www.uam.es/angeluis.villalon/cyta/fisiologiacyta3.pdf.
·
Adhesión celular, uniones celulare y matriz
extracelular [en línea] uam. Lecturas Albert cap. 19; 2008. [Fecha de acceso:
11 de septiembre de 2014] URL disponible en: http://docencia.izt.uam.mx/acbc/documentos/lecturas/alberts_cap_19.pdf.
·
Gradiente electroquímico y potencial de membrana. [en línea] [fecha de
acceso: 20 de septiembre de 2014] URL disponible en:
http://sgpwe.izt.uam.mx/files/users/uami/norm/membrana_potencial_clase.PDF
- Pilar Molist Manuel A. Pombal Manuel Megias. CITOLOGIA 4-Trafico vesicular [en línea] Depto. de Biología Funcional y Ciencias de la Salud Facultad de biología Universidad de Vigo; 2011 [fecha de acceso: 14 de septiembre de 2014] URL disponible en: http://webs.uvigo.es/mmegias/descargas/atlas-celula-05-trafico.PDF
No hay comentarios:
Publicar un comentario